Chapter 1
Distributed Cognitive Functions
A General Theoretical Framework
When approaching cognitive assessment patients in the clinic or at the bedside, it is essential to have a general structure on which to base the clinical interview and examination. The schema suggested here adopts the general neurological approach of localization followed by differential diagnosis. I have attempted to link anatomy with cognitive function, wherever possible, although the strict localization of many aspects of cognition is still far from clear. Recent functional imaging studies in normal subjects show that virtually all aspects of cognition depend upon the integrated activity of several brain regions. In a quest for clarity and brevity, I have been forced to take a simplified and often didactic approach that necessarily avoids many interesting issues and controversies in neuropsychology and behavioural neuroscience. For those wishing to read more detailed analyses of brain structure–function relationships, and of cognitive neuropsychology in general, several reference sources are suggested in ‘Selected Further Reading’ at the end of this book. Thankfully, a number of excellent textbooks have been published in the last few years.
The basic dichotomy offered here is between distributed and localized functions. The label distributed implies cognitive abilities which are not strictly localized to one lateralized brain region, as outlined in Table 1.1. Hence abnormalities of these distributed functions do not, with a few notable exceptions (such as amnesia following thalamic strokes), arise from small discrete lesions, but typically result from fairly extensive and often bilateral damage or more generalized insults of the type encountered in general medical practice. Localized functions can, in turn, be divided into those associated with the dominant, usually left, hemisphere, and those associated with the nondominant hemisphere.
In this chapter, I shall describe three broad domains of cognition that have a distributed neural basis: arousal/attention, memory, and executive function. Chapter 2 deals with the syndromes of delirium and dementia and Chapter 3 covers localized cognitive abilities. The tests mentioned in Chapters 1, 2, and 3 will be described more fully in subsequent chapters.
Table 1.1 Distributed cognitive functions
| Cognitive function | Neural basis |
|---|---|
| 1. Attention / concentration | Reticular activating system (brainstem and thalamic nuclei), and multimodal association areas (prefrontal and parietal) with right bias |
| 2. Memory | Limbic system (especially hippocampus and diencephalon) |
| 3. Higher-order executive functions and social cognition | Frontal lobes |
Arousal and Attention
Although attention to the external environments and to our own internal thought processes are clearly extremely important, it remains difficult to state the specific defining characteristics of attention. We all know what to be attentive means in ecological terms, but cognitively, attention is a complex ability. During wakefulness, an individual is bombarded with a plethora of sensory stimuli originating from the environment arriving via all of our sensory organs. In addition, we are also in a state of constant rumination with thoughts, ideas, and memories which pop into mind, often seemingly at random. Despite this, we are able to engage in specific goal-directed behaviours ranging from making breakfast or driving to work to chairing a meeting or solving a series of complex problems. Our attentional processes allow us to focus on specific parts of stimulus space and to hold other stimuli at bay, at least temporarily. We are also able to shift our attention and often to engage in two tasks at once.
There have been many attempts to characterize the various subprocesses embraced within the broad rubric of attention. For clinical purposes, the following is a useful classification of the components of attention:
A good example of the application of these processes is the ability to drive a car while conversing with a passenger under easy driving conditions, which requires both sustained and divided attention, but as soon as a more complex operation, such as merging lanes or overtaking, is required then it is usually necessary to shift attention and to focus on the operations involved with driving. The four processes described in the listed classification are all global, or superordinate, aspects of attention, which operate across sensory domains and can be contrasted to domain-specific attentional abilities (see later in this section).
Orientation, concentration, exploration, and vigilance are positive aspects of global attentive processes, while distractibility, impersistence, muddledness, and confusion reflect impaired attention. Marked impairment of attention is almost always accompanied by disorientation in time or place, or both. The clinical syndrome, which exemplifies a breakdown of global attention at processing, is the acute confusional state (sometimes called acute organic psychiatric syndrome or, more simply, delirium). Although other abnormalities are found in delirium, disordered attention is the principal and most consistent disturbance. When severe, consciousness is diminished due to depression of basic arousal processes.
Rather confusingly, the term attention is also used in the context of so-called domain-specific attention. The most well-known aspect of domain-specific attention is attention to space. Breakdown of this ability results in the spatial neglect commonly seen after right hemisphere strokes. Regions in the nondominant hemisphere, particularly the inferior parietal and prefrontal regions, have a specialized role in spatial attention, which will be discussed further under the heading of neglect (see Chapter 3).
It is important to know that wakefulness or arousal is only one aspect of attention. In states of diminished wakefulness, noxious stimulation is required to provoke a response that is typically stereotyped and non-purposeful. Patients in such a state are described as being drowsy, stuporosed, or in a coma, depending on the level of arousal deficit. In patients with diminished awareness, further testing of cognition is clearly pointless and scales such as the Glasgow Coma Scale are more appropriate assessment tools.
Applied anatomy
The maintenance of attention depends upon the interaction of two major neural systems: the ascending reticular activating system (ARAS) which exerts so-called bottom-up modulation of cortical regions and a cortical ‘top-down’ system of regulation involving limbic, parietal, and especially prefrontal cortical regions. In addition to these two domain-independent systems, there are local ‘domain-specific’ processes operating in specialized cortical regions which modulate responsivity to sounds, tactile stimuli, motion, faces, objects, words, and memories. This overall attentional matrix (in the terms of Marsel Mesulam) controls the diverse processes described earlier in this section (see Fig. 1.1).
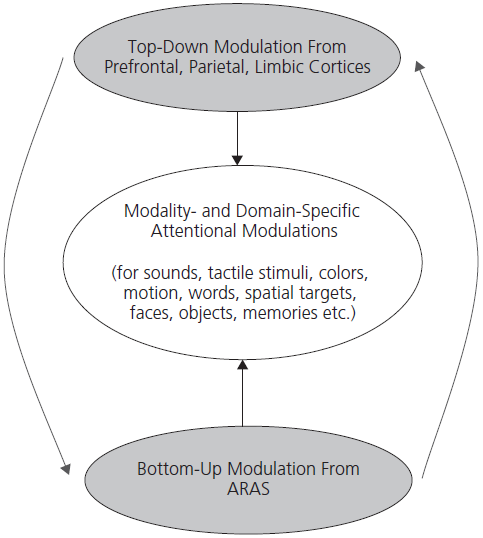
Fig. 1.1 Three compartments of attentional matrix.
Reproduced from Mesulam, M. M. (ed.) (1985). Principles of Behavioural and Cognitive Neurology (2nd edition). By permission of Oxford University Press.
Attention is therefore a collective manifestation of bottom-up, top-down, and domain-specific modulation. Disruption of the bottom-up ARAS system produces the syndrome of delirium or, if very severe, coma. Pathology involving the top-down system tends to cause less pronounced deficits of the type of inattention and distractibility seen following frontal and parietal damage as a result of traumatic brain injury or stroke.
The ARAS contains several components as shown in Fig. 1.2. Perhaps the best known is the reticulothalamic cortical pathway, which promotes and maintains cortical arousal by facilitating the transthalamic passage of sensory information towards the cortex. Acetylcholine is the major neurotransmitter in the lower, reticulothalamic component of this pathway, while excitatory amino acids (such as glutamate) are important thalamic–cortical transmitters. Other important components of the ARAS are transmitter-specific pathways originating in the brainstem or basal forebrain and projecting to the cerebral cortex. These brainstem components include the dopaminergic projections from the raphe nucleus and the noradrenergic projections from the locus coeruleus. The basal forebrain component includes the cholinergic and GABAergic pathways originating in the nucleus basalis. Bottom-up modulation of attention depends therefore upon a delicate balance of brain chemical systems. It is not surprising that diverse metabolic abnormalities produce delirium. It should be recalled also that many of these systems are involved in neurodegenerative disorders, which helps to explain the inattentiveness that occurs from an early stage in Parkinson’s disease and related syndromes, Alzheimer’s disease, and particularly in dementia with Lewy bodies.
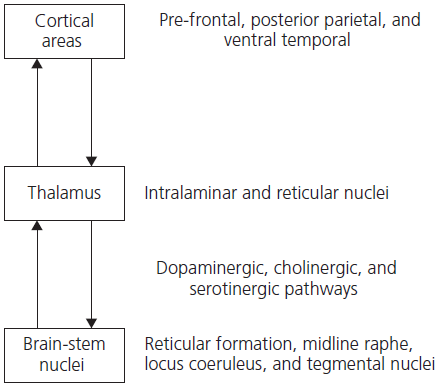
Fig. 1.2 Ascending reticular activating system (ARAS): major structures and pathways involved in normal attentional process.
The posterior parietal, limbic, and prefrontal cortices mediate top-down modulation of attention. The parietal cortex is particularly active during functional imaging studies in normal subjects engaged in tests of sustained and selective attention while the dorsolateral prefrontal cortex has a key role in divided attention. Within the limbic system, the anterior cingulate cortex plays a pivotal role in attention, as demonstrated by a series of functional imaging studies in normal subjects. Damage to this region, following bilateral anterior cerebral artery occlusion, or in association with butterfly glioma, produces a state of severe inattentiveness referred to as akinetic mutism.
The thalamus acts as a major relay station between the cortex and the ARAS. The intralaminar nuclei receive inputs from brainstem nuclei and relay information widely to the cortex. A reciprocal feedback loop from the cortex modulates these ascending pathways via the thalamus.
It can be seen from this brief description that global disorders of attention can arise from diverse pathologies involving these bottom-up and top-down modulatory systems. This disruption may result from structural damage or, more commonly, metabolic disorders and pharmacological agents, as described more fully later. Lesser degrees of attentional impairment involve specific components such as selective sustained or divided attention. For instance, impairment in selective attention appears to be particularly common in the early stages of Alzheimer’s disease, although such patients also go on to develop divided attention deficits. In dementia with Lewy bodies, there is profound impairment of attentional processing and patients with vascular dementias likewise have marked problems with attentional modulation.
Tests of attention
Memory
Introduction
Neuropsychological research in humans with focal brain lesions and in nonhuman primates following surgical lesions has shown that memory is not a single all-encompassing system. Unfortunately, a plethora of terms has arisen to describe the various subcomponents of memory. One broad distinction divides memory into that available to conscious access and reflection (called explicit or declarative memory), and those types of learned responses, such as conditioned reflexes, motor skill acquisition, and priming, which are not available for conscious reflection (called implicit or procedural memory). Explicit memory is further divided into two systems: one is responsible for the laying down and recall of personally experienced and temporally specific events or episodes, and is called episodic memory; the other type of explicit memory is responsible for our permanent store of representational knowledge of facts and concepts, objects and people, as well as words and their meaning, and is termed semantic memory. Although in linguistics and philosophy ‘semantics’ refers purely to the study of word meaning, in neuropsychology semantic memory has a wider use and applies to our general store of world knowledge. Semantic memory begins to be acquired early in life and continues to expand throughout our lifetime. It is organized conceptually, without reference to the time and context in which it was acquired. Both episodic and semantic memory are components of our long-term memory systems (see Table 1.2 and Fig. 1.3).
To illustrate this dichotomy further, consider the example of recalling the details of a conversation from earlier in the day or a dinner on holiday in Paris last year, both of which depend upon episodic memory. By contrast, knowing that Paris is the capital of France, what the word ‘dinner’ means, that a canary is a small, yellow bird more closely related to a sparrow than a penguin, and identifying a photograph of Bill Clinton all depend upon our semantic memory. The contents of episodic and semantic memory are both available to conscious access. By contrast, the acquisition of motor skills, such as learning to drive a car or play a musical instrument, does not use explicit memory, but instead relies upon the implicit memory system.
The term ‘memory disorder’ may, therefore, apply to various different types of problem. Most commonly, it is used to mean a disorder of episodic memory, that is to say, difficulty recalling personally experienced episodes from the recent past or learning new information. Disorders of episodic memory will be dealt with in more detail later in this chapter. In brief, they occur either in the context of diffuse brain injury (as part of a dementia), or as a result of selective damage to bilateral limbic structures, when the disorder is pure, spares other aspects of cognition, and is termed the ‘amnesic syndrome’.
Table 1.2 Divisions within long-term memory
| Type of material | Neural substrate | |
|---|---|---|
| Explicit | ||
| Episodic | Personally experienced episodes and events; time and context specific | Extended limbic system |
| Semantic | Vocabulary, facts, concepts, object and face knowledge; time and context specific | Polar and inferior temporal neocortex (with lateralized specialization) |
| Implicit | ||
| Procedural | Motor skills, e.g. driving, playing golf Priming Classical conditioning |
Basal ganglia Cerebral cortex Unknown, cerebellar? |
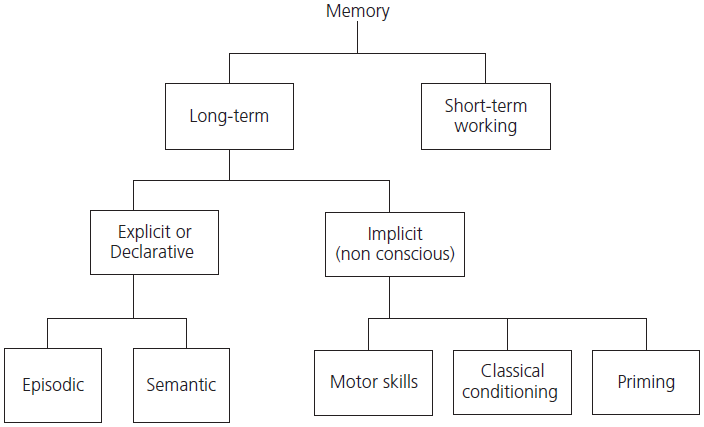
Fig. 1.3 Major subdivisions of memory.
Semantic memory loss is also an integral part of many dementing illnesses, particularly Alzheimer’s disease. Isolated impairment of semantic memory is uncommon, but occasionally occurs in the context of anterior temporal lobe damage, particularly in the syndrome of semantic dementia. Profound impairment of episodic and semantic memory occurs in survivors of herpes simplex virus encephalitis.
Short-term (working) memory
In neuropsychological terms, short-term memory is synonymous with the system of working memory responsible for the immediate recall of small amounts of verbal (as in, for example, digit span) or spatial material. It was traditionally held that for new information to enter long-term memory it must first pass through this short-term memory store. Likewise, it was believed that material recalled from long-term memory stores must first be processed by the immediate store. This simplistic serial processing model was rejected following the discovery of brain-injured patients with defective short-term memory but with completely normal ability to lay down and retrieve new longer-term memories. In addition, a number of apparently normal subjects (usually undergraduates who unwisely volunteered for psychology experiments!) have been found to have very limited short-term memory capacity. It is worth noting at this point that patients with very severe deficits in forming new memories, as in Korsakoff’s syndrome, have normal short-term memory.
There is now good evidence that there are, in fact, various subcomponents of working memory responsible for the immediate repetition of words, numbers, and melodies (called the phonological or articulatory loop) and separately for rehearsal of spatial information (the so-called visuospatial or visual sketch pad), both of which are controlled by a system known as the central executive (see Fig. 1.4). Working memory appears to function independently of, but in parallel with, longer-term memory. The central executive component of working memory is associated with the dorsolateral prefrontal lobe and is particularly important for dual-task performance, that is to say, when two tests are performed simultaneously. This aspect of working memory is really part of the larger attentional system discussed earlier.
The phonological loop depends upon perisylvian language areas in the dominant (typically left) hemisphere and the visuospatial sketch pad is associated with non-dominant parieto-occipital regions. Hence damage to widely dispersed brain regions may impair distinct components of working memory. For example, a reduced digit span is common in aphasic patients with left hemisphere lesions and is also seen in patients with frontal lobe pathology. The mechanisms underlying this deficit are different in these two instances. In the former, the phonological loop system is defective, typically producing a marked reduction in both forward and backward digit span, as well as problems in word and sentence repetition. Span reduction in association with dorsolateral prefrontal lesions is typically more modest involving particularly reverse span and, in contrast to phonological deficits, sparing word and sentence repetition. This is also a common finding in patients with frontal and subcortical dementia syndromes.
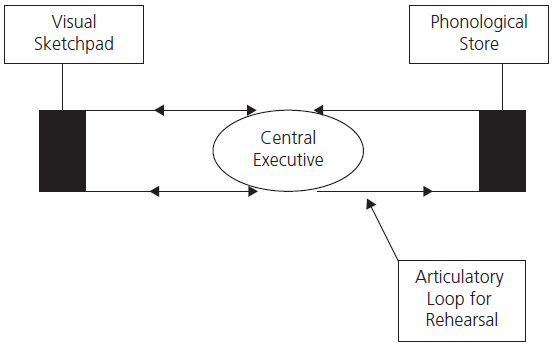
Fig. 1.4 Working memory model.
Adapted from Human memory: theory and practice, Baddeley, A.D, Copyright (© 1997) and Psychology Press, reproduced by permission of Taylor & Francis Books UK.
Clinicians use the term ‘short-term memory’ loosely to refer to recall of new material over an ill-defined short period, typically between 5 and 30 minutes, but sometimes they refer to retention over days or weeks. There is no evidence, however, either from the study of normal subjects or patients with brain disorders, to support the existence of a storage system with these temporal characteristics. As discussed earlier, the neuropsychological evidence points to there being one system that is responsible for very short (or immediate) recall of verbal or spatial material (termed working memory), and a number of longer-term systems responsible for different types of material: episodic, semantic, procedural, etc. This brief discussion highlights the controversy over what is meant by ‘short-term memory’. I have therefore avoided discussing short-term memory. When I do apply this term it is used in the neuropsychological sense to refer to immediate or working memory. In clinical practice, a much more useful distinction is between the acquisition of new information (anterograde memory) and the recall of previously learnt material (retrograde memory) since these two components may be impaired independently in different pathologies, as we shall see later in this chapter.
Tests of working memory
Episodic memory
Applied anatomy
Extensive studies of patients with acquired focal lesions as well as functional brain imaging studies in normal control subjects have established which structures are critical for the laying down and retrieval of episodic memories: the medial temporal lobe (particularly the hippocampus, subiculum, and the entorhinal cortex), the diencephalon (mamillary bodies, plus the anterior and dorsomedial nuclei of the thalamus with their interconnecting tracts), the basal forebrain nuclei (the septal nucleus, the diagonal band of Broca, and the nucleus basalis), and the retrosplenial cortex. All of these structures are bilateral with components in both hemispheres. The principal areas are connected by a number of pathways, including the fornix and the cingulate gyrus. Together these structures constitute the extended limbic system, sometimes referred to as the circuit of Papez (see Fig. 1.5). The hippocampus has traditionally been viewed as the core or central component of this system. It receives afferents from, and sends efferents to, each of the higher-order sensory association areas (visual, auditory, somatosensory, etc.) as well as polysensory areas. The internal circuitry of the hippocampus has been worked out in great detail: inputs arrive at the dentate gyrus via the perforant pathway, which sends efferents back to the association areas and to the mamillary bodies via the fornix; the dentate gyrus projects to CA3 which, in turn, projects to CA1; the latter relays to the subiculum.
Damage anywhere in the limbic system can produce memory deficits but these are often subtle and material specific. For instance, surgical removal or infarction of the left hippocampus produces a selective memory deficit for verbal material. By contrast, right-sided damage to the same structures produces a specific non-verbal memory problem (for instance, learning new faces or spatial information) which would not be apparent without detailed neuropsychological assessment. Bilateral damage to the medial temporal lobe, the diencephalon, or the basal forebrain produces a devastating and profound amnesic syndrome for both verbal and non-verbal material.
There is ongoing debate about specialization within medial temporal lobe structures in terms of recall and recognition memory. There is growing evidence that the hippocampus proper is particularly important for spatial memory and for the recall, as opposed to recognition, of newly learnt material. In addition, it has a special role for cross-modal associative learning: that is to say, learning to associate different types of sensory information such as a pattern with a particular spatial location, or a face with a name.
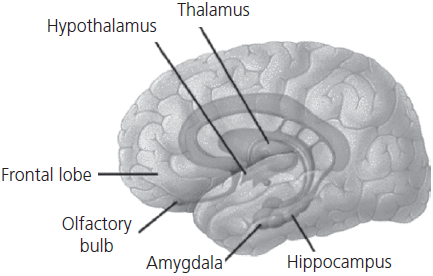
Fig. 1.5 Principal components of the limbic system concerned with episodic memory.
Evidence from patients with Alzheimer’s disease who suffer from severe impairment of episodic memory has implicated the retrosplenial cortex as playing a key role in memory processing. Recent studies in non-human primates with surgically induced lesions have emphasized the role of the parahippocampal structures, particularly the perirhinal cortex. Lesions to this area cause profound memory impairment affecting both recall and recognition. The relationship between these medial temporal regions is shown in Fig. 1.6 and an anatomical illustration is given in Fig. 1.7.
Disorders of episodic memory: transient amnesia and the amnesic syndrome
Impaired episodic memory is a feature of both delirium and dementia and in both of these disorders the memory deficit is multifactorial, with contributions from decreased attention, impaired retrieval strategies, and defective memory per se. The term amnesic syndrome should be restricted to patients with pure disorders of memory, sparing other intellectual abilities. Such disorders may be acute and transient, or chronic and generally permanent (see Table 1.3).
Transient amnesia
Transient global amnesia (TGA) is an interesting and not uncommon disorder. It typically occurs in later life with a peak around the sixth and seventh decades. Cases under the age of 40 years are exceptional. The apparent tail-off in very late life is probably due to under-reporting. The patient, who is generally in good health, suddenly becomes profoundly amnesic. Short-term (working) memory is preserved, but he or she is unable to retain any new information for more than a few seconds. The profound anterograde amnesia is accompanied by a variable retrograde memory deficit spreading back weeks, months, or even years. Patients appear disorientated and repetitively ask the same cycle of questions (for example, ‘What’s happened to me?’, ‘Why am I here?’, or ‘What day is it?’) but there is no impairment of conscious level or attention, and no language or visuospatial deficits. After a few hours, typically 4–8, the ability to lay down new memories returns gradually and the retrograde amnesia shrinks so that patients are left with a dense amnesic gap encompassing the duration of the attack, often together with the couple of hours preceding the onset. The rate of recurrence is in the order of 2–3% per year and the general prognosis is excellent. The cause of this syndrome remains unknown but it is clear that thromboembolic cerebrovascular disease does not play a part, at least in the vast majority of cases. There is an association with migraine and with stressful emotional and physical events immediately preceding the attack.
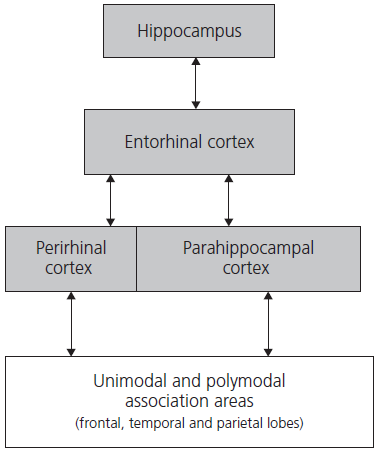
Fig. 1.6 Subregions of the medial temporal lobe.
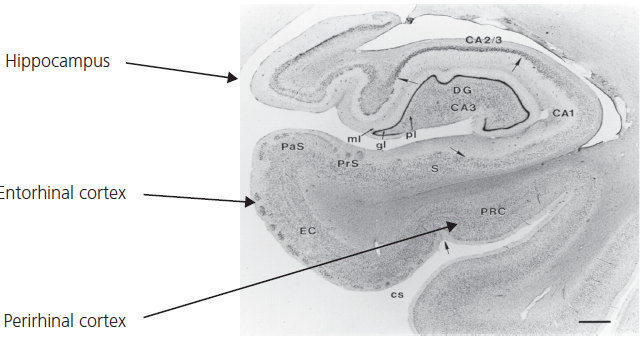
Fig. 1.7 Anatomical cross-section of hippocampus and related parahippocampal structures.
Transient epileptic amnesia (TEA) is a more recently recognized syndrome that resembles TGA. Patients are typically in the same age range as those with TGA although it can occur in younger subjects. Patients present with brief episodes of confusion and disorientation. There is repetitive questioning, as in TGA, but the duration of attacks is generally less than an hour and typically only a few minutes. Attacks often occur immediately upon waking in the morning or after a catnap. Unlike TGA, there can be partial recall of the ictus. Episodes are recurrent and may evolve into more typical complex partial seizures. Most patients with TEA also complain of large gaps in their autobiographical memory which come to light when trying to recall the details of holidays or family events undertaken during the past few years. A third characteristic of TEA is the complaint of accelerated long-term forgetting, which means that a patient’s ability to recall new information after a short delay of a few hours or days is typically within normal limits, but after a few weeks such information is lost. This can be demonstrated by testing memory for new material after a longer than usual interval. Regular electroencephalogram (EEG) recordings may be normal, but sleep EEGs usually reveal temporal lobe spike or sharp wave activity. The aetiology remains mysterious but in a few cases it may have a cerebrovascular origin. Treatment with anticonvulsants typically prevents recurrence of the TEA events but may not improve remote memory loss.
Table 1.3 Causes of episodic memory impairment
| Pure amnesia | Mixed (other accompanying cognitive deficits) | |
|---|---|---|
| Acute (transient) | Transient global amnesia Transient epileptic amnesia Closed head injury Drugs, e.g. benzodiazepines, alcohol Psychogenic (hysterical) fugues |
Delirium (see Chapter 2) |
| Chronic (persistent) | Amnesic syndrome: | Dementia (see Chapter 2) |
| 1. Hippocampal damage Herpes simplex virus encephalitis Limbic encephalitis (paraneoplastic) Anoxia Surgical removal of temporal lobes Bilateral posterior cerebral artery occlusion Closed head injury Early Alzheimer’s disease |
||
| 2. Diencephalic damage | ||
| Korsakoff’s syndrome (alcoholic and non-alcoholic) Third ventricle tumours and cysts Bilateral thalamic infarction Post-subarachnoid haemorrhage especially from anterior communicating artery aneurysms |
||
| 3. Retrosplenial damage Tumours Bleeds Alzheimer’s disease |
Acute closed head injury may cause a state very similar to TGA but there is usually very limited retrograde amnesia, and attentional processes are also typically impaired.
Hysterical or psychogenic fugue states are now rare. They occur predominantly in younger adults with a background of psychiatric problems. There is usually a recognizable precipitating life event such as bereavement or impending criminal charges. In contrast to TGA there is a profound retrograde amnesia encompassing the subject’s whole life and including loss of personal identity but sometimes without significant anterograde memory impairment.
The amnesic syndrome: defining characteristics
The amnesic syndrome: cognitive neuropsychology
There is still considerable controversy regarding the basic cognitive deficits underlying the amnesic syndrome. Short-term (working) memory, as judged by digit span or registration of a name and address, is normal. According to contemporary information-processing accounts of memory, the deficit in amnesia could be at any of the following stages:
Patients with diencephalic amnesia, as in Korsakoff’s syndrome, have problems mainly with memory encoding and, hence, lay down weak memory traces. Once information has entered into long-term stores there is little evidence of rapid decay or forgetting. Patients with Korsakoff’s syndrome also have an extensive and temporally graded retrograde amnesia. It has been argued that this remote memory deficit is due to the fact that alcoholics have been encoding weak memories for many years, because they spend their life in an alcoholinduced blur. A number of studies have refuted this suggestion, and it appears that retrieval per se is also defective in Korsakoff’s syndrome. Moreover, exactly the same picture of severe anterograde and retrograde amnesia can be seen in patients with non-alcohol-induced (for example, starvation-induced) Korsakoff’s syndrome, and in patients with bilateral thalamic infarction. Hence there seems to be dual pathology, affecting the laying down of new memories, as well as the retrieval of both new and old memories, in diencephalic amnesia.
Hippocampal amnesia, as a result of anoxia or very restricted infarction of medial temporal lobe structures, has traditionally been associated with impaired anterograde amnesia but a temporally limited retrograde amnesia: some patients are said to show deficits in recall of autobiographical memories from only the most recent 2–5 years. To explain this pattern, memory theorists argue that the hippocampus acts as a temporary ‘link’ system for newly acquired memories: the individual elements constituting a new episode, such as images of people, places, and fragments of conversation, are held to be stored in appropriate areas of sensory cortex but the hippocampal formation provides the essential link that binds the elements into a distinct episode. The hippocampus, it is argued, has limited neural space and most trivial memories fade so that space can be re-used. More significant events, which are rehearsed and refreshed by reminiscence, retelling, or perhaps even dreaming, establish more permanent connections in the cortex and gradually become independent of the hippocampal formation, a process known as consolidation.
Recent findings in patients with apparently circumscribed damage to the hippocampus have cast doubt on this model. It appears that some hippocampal patients may have a temporally extensive retrograde memory loss encompassing the whole of their life. To explain this finding, opponents of the standard consolidation model argue that memory rehearsal and refreshment leads to the formation of multiple traces in the hippocampus itself (the multiple trace model) and that the extent of retrograde amnesia depends, therefore, upon the extent of hippocampal formation damage. Hence partial damage will cause a temporally limited retrograde loss whereas more severe damage will produce extensive remote memory loss. Upholders of the standard model argue that patients with temporally extensive loss must have damage to other brain structures. This controversy remains unresolved, although recent brain activation studies in normal subjects support the multiple trace position.
It also appears that structures closely related to the hippocampus, notably the entorhinal and perirhinal cortices, have a key role in aspects of memory previously ascribed to the hippocampus. The hippocampus itself is still thought to be critical for spatial memory and for so-called cross-modal association, for instance, learning that a particular pattern or face occurred in a particular spatial location. By contrast, recognition memory depends more upon these parahippocampal structures.
Another recently recognized phenomenon in patients with amnesia due to hippocampal pathology or lesions involving other critical components of the Papez circuit is the inability to imagine possible future scenarios involving themselves.
Memory impairment is also a major and early feature of Alzheimer’s disease and other dementing disorders. In Alzheimer’s disease the situation is more complex than that in the amnestic syndrome. Each of the processes involved in long-term memory—encoding, consolidation, and retrieval—may be impaired. One of the hallmarks of early Alzheimer’s disease is a rapid forgetting of any new material, but there is also extensive retrograde amnesia, implying either problems with retrieval or a loss of stored information. Initially, short-term (working) memory may be normal but with disease progression this too becomes defective. In addition, semantic memory breaks down, so that the patient’s database of knowledge about the world progressively declines, leading to a range of deficits, including diminished vocabulary, impaired word comprehension, and difficulty in naming objects.
Frontal lobe: attention, working memory, and the temporal aspects of episodic memory
Patients with damage to the dorsolateral prefrontal cortex frequently complain of poor memory and forgetfulness, which is even more apparent to family members. Yet unlike patients with medial temporal lobe and diencephalic lesions they perform relatively well on standard memory tests: they are likely to show weak performance on word-list learning tests and recall of stories but do much better on recognition-based memory tests. They cannot, therefore, be considered as classically amnesic. This discrepancy between symptoms and test performance can be understood in terms of the difference between the processes required for effective memory use and those involved in memory encoding, storage, and consolidation. Another way of looking at this is to regard the prefrontal cortex as the conductor of the memory orchestra. Without a conductor there is chaos even although all of the players are present. In addition, the prefrontal cortex has a specific role in the temporal aspects of episodic memory. Patients with damage to this region mistake when they learnt various things, leading to a mix-up or conflation of past memories and a form of confabulation. Another more colourful form of confabulation, known as fantastic confabulation, is much rarer and occurs in the acute stages of Wernicke–Korsakoff syndrome and in patients with basal forebrain damage following subarachnoid haemorrhage and also after brain trauma. Such patients relate the occurrence of things that did not happen to them, for example, saying that they went to London last week and had tea with the Prime Minister.
Tests of anterograde episodic memory
Verbal
Non-verbal
Memory batteries
Tests of retrograde memory
Personal (autobiographical)
Semantic memory
The storage, maintenance, and retrieval of factual information and vocabulary does not, in contrast to episodic memory, depend upon the limbic system. New facts and words are assumed to be learnt in the context of an episode; but at some stage, perhaps with repeated rehearsal, they enter our fund of general knowledge. Their retrieval then becomes independent of the personal and time-tagged labels essential for recreating episodic memories. To illustrate this difference consider attending a lecture on a new topic: recall of the information is at first very dependent upon the context, and proceeds by reconstruction of the events at the time of acquisition; with repeated exposure (or retrievals) some of the information becomes part of our general store. Who can recall where and when they first learnt the meaning of the word haemorrhage, or the name of the capital of France? There is, however, recent evidence that children with early life damage to their hippocampal system (developmental amnesia) do acquire semantic knowledge even in the absence of a normally functioning episodic memory system, suggesting that, with repeated exposure, non-hippocampal parts of the temporal lobe can acquire new semantic memories.
It is important to point out that deficits of semantic memory do not simply affect word-based knowledge (such as the ability to name objects or to produce word definitions) but involve the fundamental knowledge-base underlying word knowledge. Current cognitive psychology models hypothesize that there is a central, amodal, integrative store which contains abstract representations. These abstract representations are linked to modality-specific areas that contain lexical, sound, visual, or tactile information (see Fig. 1.8). Breakdown of this central amodal store has differential effects on various modalities of input. Patients invariably show greater deficits on word-based tasks because the mapping from words to meaning is entirely arbitrary. Take, for instance, the following words: echidna, platonic, or ewer. There is no way of knowing which of these words refers to an animal whereas sounds and pictures contain inherent or deducible information.
The study of semantic memory is relatively recent, but current evidence suggests that the anterior temporal neocortex, particularly in the left hemisphere, is the key integrative region linking other, more posterior, temporal and parietal regions. Loss of semantic memory occurs as a result of extensive destruction of this anterior temporal region, often from herpes simplex virus encephalitis, or occasionally trauma or tumours. Progressive breakdown of semantic memory occurs in patients with Alzheimer’s disease, who also have a severe impairment of episodic memory. The purest and most dramatic loss of semantic memory is seen in patients with semantic dementia (a variant of frontotemporal dementia) which is associated with circumscribed polar and inferior temporal lobe atrophy.
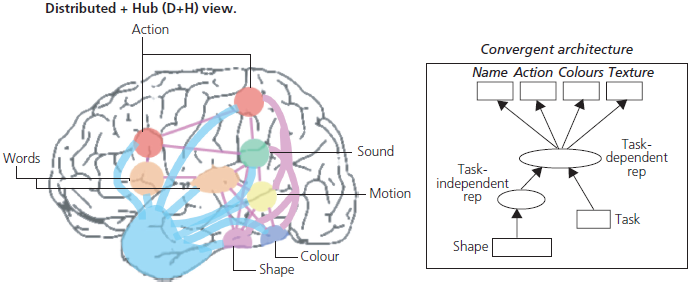
Fig. 1.8 The central amodal or ‘hub-and-spoke’ model of semantic memory. The left panel shows a computational representation with the central amodal (hub) area linking together modally specific regions (sound, speech, vision, etc.). The right panel shows an anatomical version of the model with the temporal pole as the key linkage or hub region.
Reprinted by permission from Macmillan Publishers Ltd: Nature Reviews Neuroscience, 18.1, ‘The neural and computational bases of semantic cognition’, Matthew A. Lambon Ralph, Elizabeth Jefferies, Karalyn Patterson, and Timothy T. Rogers, copyright 2016.
It seems that knowledge about different categories of things is stored separately, and in a highly organized fashion. A growing number of patients have been reported with ‘category-specific’ semantic memory loss, affecting, for instance, knowledge about living, rather than man-made, things. Patients with even finer-grained deficits, involving, for instance, fruit and vegetables and not animals, have also been reported. A loss of knowledge about living or natural things is associated with temporal lobe damage (typical after herpes simplex encephalitis) whereas the rarer converse situation (impaired knowledge about man-made things) appears to be associated with damage to frontoparietal brain regions.
Another domain of increasing interest is the domain of person knowledge. This knowledge base enables us to name a picture of Tony Blair and to access information when given the name or hearing his voice, or to match the name and face. In contrast to general semantic knowledge, the right anterior temporal lobe appears to play a key role. Loss of person knowledge occurs in Alzheimer’s disease, when it is associated with more general semantic memory breakdown, but can be seen as a relatively pure syndrome in patients with progressive right temporal lobe atrophy, which forms part of the spectrum of semantic dementia.
Disorders of semantic memory
Tests of semantic memory
See Appendix for further details.
Semantic Memory Test Battery
In Cambridge, we developed a battery of tests based on the same 64 items (half living, half man-made) that includes the following subtests: category fluency, picture naming, word-to-picture matching, picture–picture association (the Camel and Cactus Test), picture and word sorting, repetition, and generation of definitions from words (for a full description see ‘Cambridge Semantic Memory Test Battery’ in the Appendix).
A further development of this battery is the Sydney Progressive Aphasia Test Battery (SYDBAT). This battery is based upon 30 items of increasing difficulty each with a multisyllabic name. The battery comprises of the following subtests: naming, word-to-picture matching, a picture–picture association task, and word repetition (see ‘Sydney Language Battery (SYDBAT)’ in the Appendix).
Implicit memory
Both episodic and semantic memory are available to conscious access. We can reflect on both personally experienced events and our fund of knowledge of the world. However, other forms of learning occur to which we do not have conscious access. These have been termed ‘implicit’ or ‘procedural’ memory. Consider the act of learning to play a musical instrument, or learning to drive a car. Although we progressively acquire the motor skills involved in these tasks, we cannot fully explain the procedures, and improvement can only be tested practically. Another form of implicit learning goes under the term priming—in this, exposure to test stimuli improves subsequent performance, even if the subject has no conscious recollection of the initial exposure. For instance, on word-stem completion tests, subjects are first shown a list of words (for example, TRACE, BREEZE, METER, etc.), then asked to recall the list, and finally shown the initial three letters of each word (TRA, BRE, MET, etc.) and asked to complete each stem with the first word that comes to mind. Amnesic patients do well on the last task, although they have no memory of having seen the words. In a non-verbal priming test, fragmented-picture identification, subjects are shown a series of progressively less fragmented pictures of the same object, and asked to say as soon as they recognize the object. Normal subjects identify the objects much sooner (i.e. from more fragmented pictures) when re-shown the pictures. Amnesic patients demonstrate the same effect, although they deny having seen them before.
Implicit memory appears to depend neither upon the limbic system nor upon the temporal neocortex. Even patients with profound amnesia (for example, Korsakoff’s syndrome) have spared implicit memory. Current evidence points to the basal ganglia as the key region for motor learning, although priming appears to depend upon cortical areas, and the cerebellum may also be important for some classical conditioned responses.
Implicit memory is not testable at the bedside. Questioning may reveal preservation of practical skills in patients with severe impairment of explicit memory, but the main reason for its inclusion here is to make readers aware of this important and rapidly expanding area of neuropsychological investigation.
Higher-Order Cognitive Function, Personality, and Behaviour
The frontal lobes account for more than a third of the human neocortex (compared to 10% in non-human primates) and are undeniably crucial to the integrity of many aspects of ‘higher-order’ cognitive function, as well as to personality and behaviour. Damage to the prefrontal areas produces long-lasting and often devastating deficits. Yet it is notoriously difficult to define these cognitive domains accurately. Furthermore, there are no really satisfactory bedside methods of assessment. Even the neuropsychological tests traditionally described as so-called frontal lobe tasks are crude, and do not capture many of the behavioural aspects of frontal dysfunction. Here, history taking from informants and clinical observation are especially important.
Cognitive functions attributed to the frontal lobes
Adaptive behaviour:
For clinical purposes these diverse abilities can be considered under two main headings which correspond to anatomical subdivisions of the prefrontal cortex:
Executive abilities
To be effective, behaviour must be appropriate, modifiable, motivated, and free from interference and disruptive impulsive responses. If responses are to be appropriately adapted, changes in the environment need to be monitored and, if possible, anticipated. Patients with frontal lobe damage fail to anticipate changes, show poor planning ability, and do not learn from their errors. Planning is a particularly important practical function, since many complex behaviours— such as organizing a household, or holding down a job of work—require the planning and sequencing of behaviours, as well as the setting of goals. Frontal patients are especially poor at self-guided learning and goal setting. They perform normally on externally driven tasks, but are very poor at self-motivated learning. There is a striking vulnerability to interference from irrelevant stimuli, resulting in distractibility and the intrusion of unwanted responses. They also show a tendency to perseverate. Perseveration can be observed on motor tasks (such as learning a sequence of hand movements, as in the Luria Three-Step Test described in ‘Motor sequencing: the Luria Three-step Test and the Alternating Hand Movements Test’ in Chapter 5), when a compulsive repetition of movement is observed. Perseverative tendencies can also be seen on cognitive tests independent of motor activity, where subjects perseverate correct and incorrect responses. There is also an inability to shift from one task to another, and a peculiar mental stickiness described as ‘stimulus-bound’ behaviour. Many of the mentioned cognitive functions are necessary for effective problem-solving and it is, therefore, not surprising that frontal lobe damage results in severe deficits in solving problems, deducing concepts, and making analogies.
A classic test of strategy formation and shifting is the Wisconsin Card Sorting Test (WCST) which requires elements of hypothesis testing, shifting, and flexibility. The WCST forms the basis of a more sophisticated set-shifting task in the computerized CANTAB. Complex frontal executive abilities are also assessed by tests of fluid (as opposed to crystallized) intelligence such as Raven’s Progressive Matrices or Catell’s test of G. Another popular test of problem-solving is the Tower of London (originally Hanoi) task which is also available in a modified form as part of the CANTAB.
The inability to initiate and monitor cognitive strategies can also be tested by verbal fluency tasks. In the supermarket fluency test, patients are asked to list items which can be bought in a supermarket. In category fluency tests, they are asked to list as many exemplars as possible from a given category (for example, animals, fruit, vegetables, etc.) in a limited time-period, usually 1 minute. Letter fluency tests require subjects to generate as many words as possible beginning with certain letters (for instance, F, A, and S). Frontal patients show severe impoverishment in the generation of exemplars, impaired search strategies, and a tendency to repeat the same item (see ‘Initiation: verbal fluency tests’ in Chapter 5 for details of tests).
Although not involved in the laying down and storage of long-term memory traces, the frontal lobes are important for certain aspects of memory retrieval, particularly when temporal-order judgements are required (see ‘Frontal lobe: attention, working memory, and the temporal aspects of episodic memory’).
One of the components of working memory, the central executor, also depends critically upon the frontal lobes (see ‘Short-term (working) memory’). Patients with frontal lobe dysfunction may, therefore, be impaired on simple tests of short-term (working) memory (for example, reverse digit span); but they also show very marked deficits on dual-task performance tests that place heavy demands upon working memory, such as simultaneous digit span and manual tracking.
Motivation, inhibitory control, and social cognition
Patients with damage to the orbitomesial frontal lobe can suffer profound changes in personality and behaviour yet may perform normally on all the tests of executive function discussed previously. This dissociation is exemplified by the famous case of Phineas Gage who suffered major alterations in social cognition after a tamping iron penetrated his orbital frontal lobe and exited the top of his skull. Having previously been a responsible and well-adjusted, hard-working, conscientious worker he became feckless, irresponsible, indecisive, emotionally cold, and impulsive. Such cases are still seen today following road traffic accidents or surgery for orbital meningiomas, but in neurological practice, this constellation is most often seen in association with progressive frontal lobe degeneration as part of frontotemporal dementia (also known as Pick’s disease).
Considerable progress has been made over the past two decades in our understanding of these changes. The orbitomesial cortex has reciprocal connections with the amygdala, temporal pole, and insula cortex. Together, these structures constitute a critical circuit involved in social cognition and emotion processing. One important hypothesis, advocated by Antonio Demasio and colleagues, is that the orbital cortex contains so-called somatic markers, which are evoked by a range of sensory experiences and give rise to inner feelings necessary for normal mature human interactions. Patients with damage to components of this circuit may be impaired in their perception of emotions from facial expressions or from voices, whereas others can perceive emotions but have lost the appropriate valence associated with these perceptions leading to a form of acquired psychopathy.
Another theoretical advance has been the development of the concept of theory of mind, sometimes also referred to as mentalizing ability. This concept grew out of observations in individuals with autism and Asperger’s syndrome who are unable to appreciate the mental state of others. Functional brain imaging studies localize theory-of-mind abilities to the orbital and medial frontal cortices plus the superior temporal sulcus. A loss of empathy and appreciation of humour and sarcasm is thought to reflect defective theory of mind and is common in patients with frontal pathology. Another striking similarity between individuals with autism and patients with orbitomesial frontal damage is the occurrence of stereotypical ritualized behaviours akin to obsessive– compulsive disorder. Patients with frontotemporal dementia indulge in repetitive, complex patterns of ritualized behaviours such as hoarding and collecting. These behaviours are also seen in some patients with Parkinson’s disease who abuse dopaminergic therapies and are referred to in this context as punding behaviours.
Another hallmark of orbitofrontal damage is loss of inhibitory control. This results in a tendency to react immediately, and usually inappropriately, to external stimuli. Irascibility and verbal aggression are common. Male patients with orbitomesial pathology commonly make lewd jokes or exhibit sexually inappropriate behaviour.
A final aspect of cognition frequently impaired after mesial frontal lobe damage is motivation. Apathy is a very common feature and its most extreme form results in an abulic state of motionless mutism. This is rare but occurs after anterior cerebral artery occlusion or neurosurgical interventions. Less profound states of apathy are very common in Alzheimer’s disease, frontotemporal dementia, and the Parkinsonian syndromes, especially progressive supranuclear palsy.
Alterations in food preference, mainly towards sweet foods, and satiety with a tendency to overeat are also common features of frontotemporal dementia and again reflect disruption of circuits involving the orbital frontal cortex, the amygdala, and the gustatory cortex in the insula.
Frontal lobes: applied anatomy
The frontal lobes proper can be subdivided into five major areas (see Fig. 1.9):
In keeping with its role as the chief executive officer of intellectual function, the prefrontal cortex is richly connected to virtually all other subordinate cortical and subcortical structures. It receives inputs from all unimodal association areas (visual, auditory, haptic, olfactory) and the other multimodal association areas (i.e. the posterior parietal and ventral temporal lobes), as well as from the limbic structures. Major afferent projections arise from the dorsomedial nucleus of the thalamus and basal ganglia. This accounts for the ‘frontal lobe’ or dysexecutive deficits that typically occur in the subcortical dementia syndrome associated with basal ganglia disorders (for example, Huntington’s disease, progressive supranuclear palsy, and Parkinson’s disease), and in lesions of the thalamus.
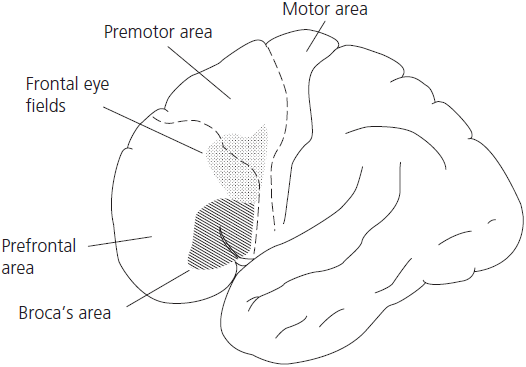
Fig. 1.9 Dorsolateral frontal cortex with functional areas.
The orbitomesial frontal cortex is richly interconnected with the polar parts of the temporal lobe and the amygdala as discussed earlier.
Disorders of frontal lobe function
Degenerative
Vascular
Structural
Deafferentation from basal ganglia disorders
Examples include:
Tests of frontal lobe function
See Chapter 4 and Appendix for further details.